Esta es la primera de una serie de entregas en el campo de la biofísica.
En ellas discutiremos despacito y con buena letra el mecanismo por el que se expresa un gen desde el punto de vista físico, haciendo especial hincapié en la aleatoriedad inherente del proceso.
Vamos a ello…

El ADN se copia a ARN y el ARN se traduce a proteínas. Estos son los pasos básicos de la expresión de un gen en proteínas.
De ADN a ARN y de ARN a Proteína
Esto es parte de lo que se conoce como dogma central de la biología molecular.
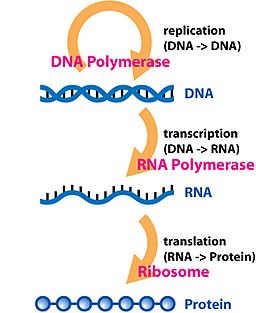
Este establece que la información genética está codificada en los genes.
Un gen es una secuencia de ADN que contiene información relevante. Originalmente se pensaba que existía esta relación:
Un gen codifica una proteína
Hoy día sabemos que esto no es correcto y que la información genética es mucho más amplia que esto. Sin embargo, esta aseveración no deja de ser correcta.
Lo que estamos seguros es que hay proteínas que están codificadas por un gen.
El mecanismo para descodificar un gen es el siguiente:
- Tenemos una secuencia de ADN (que estará en algún lugar del citoplasma para células procariotas, las bacterías por ejemplo, o en el núcleo para células eucariotas, por ejemplo células vegetales o animales.) que codifica una proteína específica.
- El ADN se activa, ya veremos cómo en las entradas de biología, y empieza a transcribirse a ARN. El ARN es una clase de material genético que comparte similitudes con el ADN en términos de composición química. Se diferencian en que el ARN suele ser una única cadena y el ADN son dos cadenas de nucleótidos en forma de doble helice. El ARN resultado de la transcripción se conoce como ARN mensajero.
- El ARN mensajero se transporta a sitios de la célula donde hay ribosomas. Estos ribosomas son ensamblajes proteínicos esencialmente que tienen la capacidad de que pueden “leer” el ARN y hacer la traducción a proteínas. Pedestremente van leyendo en el ARN mensajero qué aminoácidos hay que ir insertando para formar la proteína.
Este mecanismo es muy complejo (en términos de participación de distintas encimas y complejos moleculares) pero con esta breve descripción tendremos suficiente para entender lo que nos interesa ahora, desde el punto de vista físico, de este proceso.
Se nota que eres físico…
En física usualmente se reduce el problema a tratar a los elementos esenciales que capturan su naturaleza para poder modelizarlo matemáticamente.
Esta ocasión no íbamos a ser menos.
Un físico ve el paso de ADN a ARN mensajero (ARNm) como algo que será más o menos complicado de conseguir pero que se realiza con una determinada velocidad (de reacción)  :
:
:![ADN \xrightarrow[]{s} ARNm](http://s0.wp.com/latex.php?latex=ADN+%5Cxrightarrow%5B%5D%7Bs%7D+ARNm&bg=ffffff&fg=333333&s=0 "ADN \xrightarrow[]{s} ARNm")
donde  , es decir una velocidad constante.
, es decir una velocidad constante.
, es decir una velocidad constante.
Esto es así porque sabemos que este procedimiento está mediado por un complejo entramado de enzimas que consiguen la estabilidad del proceso y el resultado neto es una reacción que se produce a velocidad constante, produciéndose s moléculas/hora (todos los elementos necesarios para la transcripción se suponen presentes y abundantes).
Un químico nos diría que esto es una reacción de orden cero.
En el siguiente paso tendremos una segunda reacción en la que se produce proteína (P) a partir del ARNm.
Es importante notar que el ARNm no se consume en la reacción.
Es decir, después de traducir un ARNm sigue estando igual que al principio.
El orden de esta reacción es uno, lo que significa que la velocidad de generación de la proteína es proporcional a la concentración de ARN, con constante de proporcionalidad k:
![ARNm \xrightarrow[]{k} ARNm+P](http://s0.wp.com/latex.php?latex=ARNm+%5Cxrightarrow%5B%5D%7Bk%7D+ARNm%2BP&bg=ffffff&fg=333333&s=0 "ARNm \xrightarrow[]{k} ARNm+P")
Y la velocidad de este paso será: ![v_2=k\left[ARNm\right]](http://s0.wp.com/latex.php?latex=v_2%3Dk%5Cleft%5BARNm%5Cright%5D&bg=ffffff&fg=333333&s=0 "v_2=k\left[ARNm\right]") .
.
.
Aquí tenemos que explicar algo. Para empezar, ![[ARNm]](http://s0.wp.com/latex.php?latex=%5BARNm%5D&bg=ffffff&fg=333333&s=0 "[ARNm]") denota la concentración de ARNm presente en la reacción inicialmente. Dado que lo único que tenemos al principio, cuando se inicia la reacción, es ARNm y no proteína es lógico que esta velocidad de reacción sólo dependa de la concentración del primero.
denota la concentración de ARNm presente en la reacción inicialmente. Dado que lo único que tenemos al principio, cuando se inicia la reacción, es ARNm y no proteína es lógico que esta velocidad de reacción sólo dependa de la concentración del primero.
denota la concentración de ARNm presente en la reacción inicialmente. Dado que lo único que tenemos al principio, cuando se inicia la reacción, es ARNm y no proteína es lógico que esta velocidad de reacción sólo dependa de la concentración del primero.
Es evidente que tanto la concentración de ARNm como de proteína dependerá del tiempo.
Bueno, ya tenemos controlado como se forma la proteína, pero hay que considerar que algo en el proceso puede ir mal y que la proteína o el ARNm se degrade y ya no sean útiles.
De hecho, aparte de accidentes existen procesos de degradación de ARN y proteínas que son esenciales para mantener el equilibrio y la viabilidad celular.
Entonces además de las dos reacciones anteriores de producción de macromoléculas, debemos considerar las correspondientes reacciones de degradación, ambas de orden uno:
![ARN \xrightarrow[]{\delta_m} \emptyset](http://s0.wp.com/latex.php?latex=ARN+%5Cxrightarrow%5B%5D%7B%5Cdelta_m%7D+%5Cemptyset&bg=ffffff&fg=333333&s=0 "ARN \xrightarrow[]{\delta_m} \emptyset")
![P \xrightarrow[]{\delta_p} \emptyset](http://s0.wp.com/latex.php?latex=P+%5Cxrightarrow%5B%5D%7B%5Cdelta_p%7D+%5Cemptyset&bg=ffffff&fg=333333&s=0 "P \xrightarrow[]{\delta_p} \emptyset")
Con su correspondiente velocidad ![v_3=\delta_m\left[ARNm\right]](http://s0.wp.com/latex.php?latex=v_3%3D%5Cdelta_m%5Cleft%5BARNm%5Cright%5D&bg=ffffff&fg=333333&s=0 "v_3=\delta_m\left[ARNm\right]") y
y ![v_4=\delta_p [P]](http://s0.wp.com/latex.php?latex=v_4%3D%5Cdelta_p+%5BP%5D&bg=ffffff&fg=333333&s=0 "v_4=\delta_p [P]") cada una.
cada una.
y cada una.
Por tanto, para cada una de las macromoléculas consideradas tenemos una reacción de generación y otra de desaparición.
Ahora se trata de intentar predecir cuál será la cantidad presente de cada una de ellas transcurrido un cierto tiempo.
Solución: El enfoque determinista
La manera más intuitiva de resolver el problema es: ¡eh! conozco el ritmo al que se produce y se destruye cada macromolécula por lo que la variación en la cantidad (que en matemáticas es la derivada respecto al tiempo) será lo que aparece menos lo que desaparece.
Así, para cada una de ellas:
![\dfrac{d\left[ARNm\right]}{dt}=s-\delta_m\left[ARNm\right]](http://s0.wp.com/latex.php?latex=%5Cdfrac%7Bd%5Cleft%5BARNm%5Cright%5D%7D%7Bdt%7D%3Ds-%5Cdelta_m%5Cleft%5BARNm%5Cright%5D&bg=ffffff&fg=333333&s=0 "\dfrac{d\left[ARNm\right]}{dt}=s-\delta_m\left[ARNm\right]")
![\dfrac{d\left[P\right]}{dt}=k\left[ARNm\right]-\delta_p\left[P\right]](http://s0.wp.com/latex.php?latex=%5Cdfrac%7Bd%5Cleft%5BP%5Cright%5D%7D%7Bdt%7D%3Dk%5Cleft%5BARNm%5Cright%5D-%5Cdelta_p%5Cleft%5BP%5Cright%5D&bg=ffffff&fg=333333&s=0 "\dfrac{d\left[P\right]}{dt}=k\left[ARNm\right]-\delta_p\left[P\right]")
Aquí tenemos un problema divertido, tenemos dos ecuaciones donde para resolver una de ellas tenemos que conocer la solución de la otra y viceversa.
Es lo que se conoce como ecuaciones diferenciales acopladas.
No entraré en la forma de resolución, que es un tanto aburrida por técnica, así que presentaré las soluciones y una gráfica que ayudará a entender dichas soluciones:
![[ARNm]=\dfrac{s}{\delta_m}\left(1-e^{-\delta_m t}\right)](http://s0.wp.com/latex.php?latex=%5BARNm%5D%3D%5Cdfrac%7Bs%7D%7B%5Cdelta_m%7D%5Cleft%281-e%5E%7B-%5Cdelta_m+t%7D%5Cright%29&bg=ffffff&fg=333333&s=0 "[ARNm]=\dfrac{s}{\delta_m}\left(1-e^{-\delta_m t}\right)")
![[P]=\dfrac{ks}{\delta_p \delta_m}+\dfrac{ks}{\delta_m(\delta_m - \delta_p)}e^{-\delta_m t}+\dfrac{ks}{\delta_p (\delta_p - \delta_m)}e^{-\delta_p t}](http://s0.wp.com/latex.php?latex=%5BP%5D%3D%5Cdfrac%7Bks%7D%7B%5Cdelta_p+%5Cdelta_m%7D%2B%5Cdfrac%7Bks%7D%7B%5Cdelta_m%28%5Cdelta_m+-+%5Cdelta_p%29%7De%5E%7B-%5Cdelta_m+t%7D%2B%5Cdfrac%7Bks%7D%7B%5Cdelta_p+%28%5Cdelta_p+-+%5Cdelta_m%29%7De%5E%7B-%5Cdelta_p+t%7D&bg=ffffff&fg=333333&s=0 "[P]=\dfrac{ks}{\delta_p \delta_m}+\dfrac{ks}{\delta_m(\delta_m - \delta_p)}e^{-\delta_m t}+\dfrac{ks}{\delta_p (\delta_p - \delta_m)}e^{-\delta_p t}")
Donde se ha impuesto la condición de que a tiempo inicial la cantidad de ARN y proteína es nula (a t = 0, [ARN] = [P] = 0).
Estas expresiones pueden tener un aspecto más o menos complejo, pero lo único que nos interesa es lo que significan: la cantidad de moléculas presentes varía de la siguiente manera (para s = 10000, k = 3,  = 10,
= 10,  = 2 ):
= 2 ):
= 10, = 2 ):
Vemos como la concentración de ARNm crece hasta hacerse constante y la de proteína aumenta conforme pasa el tiempo.
Problemas de los modelos deterministas
Los modelos deterministas no son siempre válidos.
El primer problema que se puede encontrar es que las ecuaciones anteriores están planteadas para variables continuas, no discretas.
Es decir, las ecuaciones predicen que tras media hora habrá 812,7528… proteínas, mientras que nosotros sabemos que es posible que haya 812 u 813, pero nunca una cantidad decimal.
Para que una reacción se produzca es necesario que choquen varias moléculas
")
(reactivos) y que lo hagan con suficiente energía como para que éstas se recombinen y originen los productos (choque efectivo).
Por desgracia no nos es posible conocer con exactitud cuántos choques efectivos se producen en cada intervalo de tiempo, ya que para esto necesitaríamos saber con absoluta precisión la posición y velocidad de todas y cada una de las partículas en un instante. Nos encontramos entonces con que hay un cierto grado de aleatoriedad en el número de moléculas que reaccionan en un intervalo dado.
En el enfoque determinista este problema se resuelve tomando el valor medio de la cantidad de choques efectivos, siendo este valor medio la velocidad de reacción de la que ya hemos hablado.
La pega de trabajar con promedios es que no se considera cómo de alejado puede estar un valor puntual del promedio.
Si el valor medio de choques efectivos es 100.000 y para cuatro casos concretos se tiene que los valores reales son 100.002, 100.000, 100.003 y 99.998 podemos afirmar que tomar el valor promedio es una muy buena aproximación.
Sin embargo si para ese mismo valor medio se tienen los valores 106.324, 99.871, 96.048 y 97.628 tenemos motivos para preocuparnos.
Con vistas a solucionar los dos principales problemas del enfoque determinista (uso de variables continuas y no contemplación de aleatoriedad) se emplea el enfoque estocástico para predecir el comportamiento del sistema.
En la siguiente entrega pasaremos a describirlo.
Nos seguimos leyendo… Cuentos cuánticos.